能誘導強排斥反應並參與免疫調節的基因組。
20世紀初,E.E.泰澤發現近交系小鼠的腫瘤移植到同一近交系小鼠體內後能夠生長,而移植到另一近交系小鼠體內則遭排斥。隨後P.B.梅達沃發現正常組織移植也出現類似的現象,認為移植排斥是一種免疫反應。進一步研究表明,在遺傳性不同的個體間進行組織移植,細胞表面的同種異型抗原誘導瞭針對移植物的免疫反應。G.D.斯內爾(1948)將決定移植物是否被排斥的抗原稱為組織相容性抗原(H抗原),控控制它表達的基因稱為組織相容性基因(H基因)。他隨後發現有一個H基因能誘導強移植排斥反應,稱之為主要組織相容性基因,後來發現它是由多個基因組成,並且位於同一染色體片段上,因此改稱之為主要組織相容性復合體,而將那些誘導弱移植排斥反應的H基因稱為非MHC基因或次要組織相容性基因。
1958年J.多塞發現多次輸血者和多產婦血清中含有凝集人白細胞的抗體,其特異性不同於紅細胞凝集抗體。隨後的研究發現人類白細胞抗原 (HLA)是一個極為復雜的遺傳系統的產物。這一遺傳系統稱為人類白細胞抗原復合體,就是人的主要組織相容性復合體。
60年代報道瞭對白血病病毒的抵抗力依賴於小鼠的H-2復合體。隨後又發現人的一定HLA抗原的存在和對某些疾病的易感性間存在著驚人的相關性。這導致人們對MHC生物效應的廣泛研究,取得關於免疫應答基因(Ir基因)和免疫細胞間相互作用的MHC限制性等的重大進展。
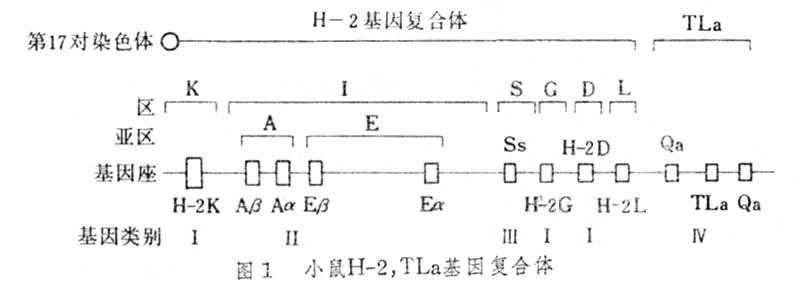
H-2基因復合體 小鼠主要組織相容性抗原受第17對常染色體 H-2基因群支配,可分為六個主要區,即K、I、S、G、D和L區。I區又可分為兩個亞區,即I-A和I-E亞區。每個區或亞區至少包括一個基因座,分別稱為H-2K座、H-2D 座、S座、H-2G座。I-A亞區含有Aα、Aβ二個座,I-E亞區含有Eα 和Eβ二個座。它們的功能各不相同,K區與D區基因可編碼 H-2抗原系,I區基因編碼的分子稱為I區相關抗原或Ia抗原系(Ia),S區基因可編碼血清因子和補體成分,G區基因與血型抗原有關。1981年J.克萊因將MHC基因座按功能分為四類(圖1)。Ⅰ類座包括K、D和L座,其編碼的分子稱為Ⅰ類分子(即K、D和L分子);Ⅱ類座,其編碼的分子稱為Ⅱ類分子(Ia分子);Ⅲ類座,其編碼的分子稱為Ⅲ類分子(血清因子和補體分子);Ⅳ類座,位於D座右側,是否屬於H-2復合體尚未確定,但與H-2連鎖,它包括TLa座和 Qa座,其編碼的分子稱為Ⅳ類分子(TL分子和Qa分子)。
HLA 基因復合體 1958年法國J.B.G.J.多塞首先在多次受血者的血清中發現能凝集人白細胞的抗異型白細胞抗原的抗體,並鑒定瞭第一個人白細胞抗原,定名為Mac(這是三個受檢者名稱的第一個字母)。其後近10年的研究證明,人組織相容性抗原也受多基因座控制,該基因座稱為人主要組織相容性基因復合體(HLA復合體)。HLA復合體定位於第6對染色體短臂上,由4個基因區(HLA-A、B、C及HLA-D區)組成(圖2)。D區相當於小鼠的Ⅰ區,由3個亞區,即HLA-DP、DQ和DR組成。HLA復合體也分為三類基因,Ⅰ類基因包括HLA-A、B、C三個座;Ⅱ類基因包括HLA-DR、DQ、DP三個座,每個基因座都表現為復等位基因,它們可編碼 HLA抗原分子;Ⅲ類基因可編碼部分補體分子。新發現屬Ⅰ類基因的 HLA-E座和屬於Ⅱ類基因的HLA-DZ、DO、DX等新出現的基因座,其定位及功能尚待進一步研究。

MHC的一個重要特征就是有高度的多態性,即在同一種屬的群體中,同一個基因座可表達二種以上乃至幾十種的基因產物。這是由於同一基因座存在著復等位基因。例如,1987年第10次國際組織相容性會議曾認為HLA-A座可表達24種抗原特異性,B座有52個,C座有11個,Dw有26個,DR座有20個,DQ座有9個,DP座有6個,共148個型別。這樣可組成107種以上的單倍型,因此在無關人群中要找到HLA完全相同的個體,幾乎是不可能的。HLA多態性為器官移植尋找合適的配型帶來瞭很大困難,但在自然狀態下,這可能是高等生物抵禦病原體的一種適應性表現。
MHCⅠ類分子(HLA-A、B、C分子和H-2K/D分子)由兩條肽鏈組成,一條為多態性糖蛋白,分子量為44000,由MHC編碼稱為重鏈(H)。另一條鏈為非多態性的β2-微球蛋白,分子量為12000,在人由第15對染色體編碼,在小鼠由第2對染色體編碼,稱為輕鏈,以非共價鍵與H鏈結合,整個分子通過H鏈固定於細胞膜上。H鏈可分為三個區,自分子的氨基末端開始,依次是細胞外親水區,穿膜的疏水區和胞內的親水區。胞外區可分為三個功能區,每個功能區可包含約90個氨基酸殘基,N-末端功能區具有寡糖側鏈的附著部位,第二、三功能區各有一個二硫鍵環。抗原的多態性主要由這兩個功能區的結構決定。胞內的親水區能被磷酸化,這種反應可能與信號傳遞有關。
在小鼠的研究表明Ⅰ類分子見於除滋養層和絨毛膜外的組織,H-2K和H-2D抗原可存在於所有組織細胞表面,在小鼠胚胎第四天就可出現H-2抗原,在人胚胎組織也可發現HLA抗原。MHC的抗原密度依細胞種類而有顯著不同,淋巴細胞密度最高,其次為肝、腎、骨骼肌和腦細胞。小鼠紅細胞上有Ⅰ類抗原,而人紅細胞上很難檢測出Ⅰ類抗原。
MHCⅡ類分子(HLA-DR,DQ,DP分子及小鼠I-A、I-E分子)亦由兩條大小不同的糖蛋白鏈組成,一條為多態性β鏈,分子量為33000,另一條為非多態性α 鏈,分子量為25000,亦為非共價結合,兩條鏈都由MHC編碼。每條鏈都由三個區組成,即胞外親水區、穿膜疏水區和胞內親水區。α 鏈胞外親水區有兩個功能區,都有一個寡糖附著部位,第二個功能區有一個鏈內二硫鍵,其胞內親水區可被磷酸化。β鏈胞外親水區也有兩個功能區,都有鏈內二硫鍵。分析瞭抗原性不同的Ⅱ類分子的氨基酸序列,發現α 鏈都很相似,而β鏈則高度不同,這表明Ⅱ類抗原分子的多態性是由β鏈決定的。
Ⅱ類抗原分子的細胞分佈與Ⅰ類抗原明顯不同,後者可存在於多種組織細胞上,而前者隻存在於少數種類的細胞,主要存在於B淋巴細胞,其次為巨噬細胞、血中單核細胞,上皮中的朗格漢斯氏細胞以及精子細胞等。不存在於靜止狀態的T細胞中,但可出現於活化T細胞上。
MHC的生理意義 MHC抗原最初是作為移植抗原而被發現的,是引起移植排斥的主要抗原系統。這種抗原不合,即可引起受體的免疫應答,排斥移植的供體組織。70年代後證明MHC分子還具有重要的免疫生理功能。
MHC分子在免疫應答過程中參與抗原識別。70年代R.M.津克納澤爾等在小鼠實驗中發現殺傷 T細胞在殺傷感染病毒的靶細胞時,隻能殺傷同系感染靶細胞,而對不同系的感染靶細胞則無殺傷作用,稱這種現象為遺傳限制性。隨後證明殺傷T細胞與靶細胞的MHC必需一致才有殺傷作用,因此又稱此現象為MHC限制性。這揭示瞭MHC在T細胞識別異種抗原中的作用。進一步的研究證明T4+的T細胞在識別異種抗原時受MHCⅡ類分子限制,而T8+的T細胞識別異種抗原時受MHCⅠ類分子限制。這種限制性的機制是:T細胞通過其抗原識別受體,可同時識別異種抗原決定簇和自己MHC分子形成的新的復合抗原決定簇。
人們還發現外周血B細胞和單核細胞等非T細胞在體外能誘導某些自身反應性T細胞發生增殖反應,稱這種現象為自身混合淋巴細胞反應(AMLR),並證明這是由非T細胞上MHCⅡ類抗原引起的。這種自身反應性T細胞在體內可能具有增強或抑制免疫功能的作用,借以維持機體的免疫穩定性,因此MHC分子也參予免疫調節作用。
研究證明,MHC分子對T細胞在胸腺內的分化成熟過程也起重要作用。體外研究發現:去除胸腺中MHCⅡ類抗原陽性的基質細胞,則T4+T細胞的發育受阻,在胸腺培養細胞中加入抗MHCⅡ類抗原的單克隆抗體,也能阻止T4+T細胞的發育。目前認為MHC分子在T細胞自身耐受的形成和T細胞庫的產生中都起著重要作用。
MHC與疾病 研究發現許多疾病與某些HLA等位基因或HLA單倍型確實呈現明顯的相關性。與HLA抗原相關的疾病有幾個應當註意的特點:病因和病理生理機理未明,以遺傳模式分佈但為弱的外顯率;與免疫異常相關;對生殖影響很少或沒有影響。
可用群體和傢系研究來證實 HLA復合體內標記基因與各種疾病狀態的相關性。因為群體研究易於進行,所以有關資料多來源於這種研究。具體疾病與某些具體HLA抗原的相關性通過計算相對風險性(RR)來定量,相對風險性可表述為具有疾病相關 HLA抗原的個體與缺乏這種抗原的個體相比發生該種疾病的機會。RR越高,在病人群體中該抗原的頻率越高。以具有 HLA-B27的強直性脊椎炎病人為例。美國患該病的高加索人種90%具有HLA-B27,美國高加索人種對照者的具有率接近9%。RR=(90×91)÷(10×9)=91。因此,HLA-B27陽性個體發生該病的風險性是 HLA-B27陰性個體的91倍。因為不同種族之間某種抗原的頻率通常有明顯的不同。所以有必要在同種族中比較病人和對照者。例如,HLA-B27見於48%的患強直性脊椎炎的美國黑人,隻見於2%美國黑人對照者,則RR=45。
已經發現很多疾病與某種抗原相關。HLA與疾病的相關性可分為統計學上較為肯定的相關性、可能相關性及潛在相關性幾組。具肯定相關性的有:強直性脊椎炎(抗原為B27)、賴特爾氏綜合征(B27)、急性前葡萄膜炎(B27)、青少年風濕性關節炎(B27)、乳糜瀉(B8)、突眼性甲狀腺腫(B8,DW3)、重癥肌無力(B8,DR3)、皰疹性皮炎(B8)、慢性活動性肝炎(BW6,DR8)、青少年糖尿病(B8,DR3)、多發性硬化(B7,DR2)。具可能相關性的有:系統性紅斑狼瘡(DR3)、天皰瘡(B13)、自身溶血性貧血(A3)、脊髓灰質炎(A3,B7)、貝赫切特氏病(B5)。具有潛在相關性的有:急性淋巴細胞白血病(A2,B12)、慢性腎小球腎炎(A2)、麻風(B14)、霍奇金氏病(A1,B8)。
為瞭解釋HLA與疾病的相關性,已提出多種假說。①結合假說:或稱受體假說 HLA抗原為細胞膜的特異受體,可與致病因子(如病毒)結合,引起免疫性或非免疫性組織損傷。②連鎖基因假說:HLA抗原伴隨具體疾病,但其本身並不直接涉及致病作用,而隻是與具體疾病的易感基因相連鎖。③分子擬態假說:某些病原體(如病毒)抗原在分子結構上與特定HLA抗原相似,因此可形成交叉耐受,機體不能產生免疫攻擊,以終止疾病過程。另一相反的觀點認為機體對病原體可發生強的免疫應答,由於病原體與HLA抗原的相似性,免疫應答同時針對外來抗原和HLA抗原,這種自身免疫應答導致疾病。某些微生物與某些 HLA抗原間的交叉反應性的資料支持這種理論。④HLA基因與Ir基因連鎖:Ir基因控制寄主對病毒或其他抗原的免疫反應性。人的Ir基因就是HLA復合體的D區基因。特定的 HLA-D區基因型可能導致特定的異常免疫反應性。這種異常免疫反應性將導致對某些疾病的易感性或抵抗性。
在不同的HLA-疾病相關中,可有不同的機理發揮作用,並且疾病的發生可能不止一種機理發揮作用。