植物從個體發生到自然死亡所經歷的全部過程,或植物由卵、孢子或其他初生階段,經過發育,一直到自然死亡的所有變化過程。
植物生活史全部變化過程,往往構成一個迴圈,稱為生活環。如上一代的卵,經過受精,發育成胚胎,長成個體,又再產生卵;或種子植物,從前一代的種子,到後一代的種子,構成的迴圈,都是生活環。
生活史和生活環的意義極其近似,有人認為沒有區別,有人認為生活史強強調某一類別的植物從初生到死亡的歷史或過程,生活環往往用來指某一種具體植物個體發育變化的過程。
基本類型 細菌、藍藻及綠藻中單細胞的原球藻等,隻有細胞分裂繁殖,沒有有性生殖。它們的生活史是最簡單的。
大多數種類的植物都有有性生殖,生活史比較復雜。依據生活史中有幾種類型的植物體,體細胞為單倍或二倍染色體,以及有無世代交替,植物的生活史可分為兩種基本類型。
單體型生活史生活史中隻有一種類型的個體,沒有世代交替的現象。依據體細胞為單倍或二倍染色體,又分兩種類型:①單體型單倍體生活史。生活史中僅有一種類型的植物體,體細胞為單倍體(n),無世代交替現象,合子在萌發前先行減數分裂。這種生活史,見於低等的藻類和菌類,如衣藻(圖1)和黑根黴屬。②單體型二倍體生活史。生活史中僅有一種類型的植物體,體細胞為二倍體(2n),沒有世代交替現象,減數分裂發生在配子形成過程中。這種生活史見於藻類管藻目、矽藻綱、褐藻綱的墨角藻(圖2)、馬尾藻等。
 圖1 單體型單倍體生活史——衣藻
圖1 單體型單倍體生活史——衣藻
1 四個雙鞭毛單倍體細胞,能反復細胞分裂,行無性繁殖,產生新個體 2 相應的細胞配對,行有性生殖 3 合子,二倍體,有4條鞭毛,能遊動 4 休眠孢子 5 休眠孢子經過減數分裂,產生4個細胞
 圖2 單體型二倍體生活史——墨角藻
圖2 單體型二倍體生活史——墨角藻
1 精子 2 卵 3 合子 4 植物體 5 生殖窩 6 卵囊 7 精囊 8 卵囊細胞分裂 9 精囊細胞分裂 10 卵囊放出卵 11 精囊放出精子
雙單體型生活史 生活史中有兩種植物體:一為二倍體,即產生孢子的孢子體,體細胞為二倍體(2n);一為單倍本,即產生配子的配子體,體細胞為單倍體(n),有世代交替現象〔見世代交替(植物)〕。這種類型在植物界中最為普遍,依據兩種個體的形態、顯著性、大小、生活期長短,以及能否獨立生活,又可分為四種類型:①等世代型。孢子體和配子體形態、大小完全一致,都有獨立生活機能,隻有體細胞一為二倍體(2n),一為單倍體(n);普遍見於藻類植物的綠藻綱,剛毛藻目的綱毛藻,石蓴目的石蓴(圖3)、苔,褐藻綱、網地藻目的網地藻,水雲目的水雲等。在低等植物中,紅藻綱和一些真菌類植物的生活史很復雜。如紅藻中的多管藻屬生活史的特點,有兩個世代,三個獨立的植物體,即雌配子體(n)、雄配子體(n)和四分孢子體(2n)(圖4)。②不等世代型。孢子體小於配子體並始終寄生在配子體上,這種類型見於所有苔蘚植物(圖5)。③不等世代型。孢子體大於配子體,兩種個體都能獨立生活,這種生活史見於藻類植物的褐藻綱海帶目的海帶(圖6),以及所有的蕨類植物(圖7)。④不等世代型。配子體小於孢子體,始終寄生於孢子體中,由於孢子體極為發達,分根、莖、葉,而配子體高度退化,因此,配子體靠孢子體周密保護和供應營養,這種生活史見於所有的種子植物(圖8、圖9)。
 圖3 雙單體型的等世代型生活史——石蓴
圖3 雙單體型的等世代型生活史——石蓴
1 遊泳孢子 2 配子體 3 配子體行無性生殖,產生遊泳孢子,直接長成配子體 4 配子體行有性生殖,產生雙鞭毛的配子 5 配子配對 6 合子 7 孢子體
 圖4 多管藻屬的生活史
圖4 多管藻屬的生活史
1 四分孢子,由四分孢子囊中的分孢子母細胞減數分裂長成 2 雄配子體 3 雌配子體 4 精囊 5 卵囊(果孢),一端具長的受精絲 6 精細胞 7 精細胞接觸受精絲,精核進入卵囊 8 精核與卵在卵囊中結合成合子 9 囊果,其中生長二倍體的果孢子世代的果孢子體 10 果孢子 11 孢子體 12 四分孢子體
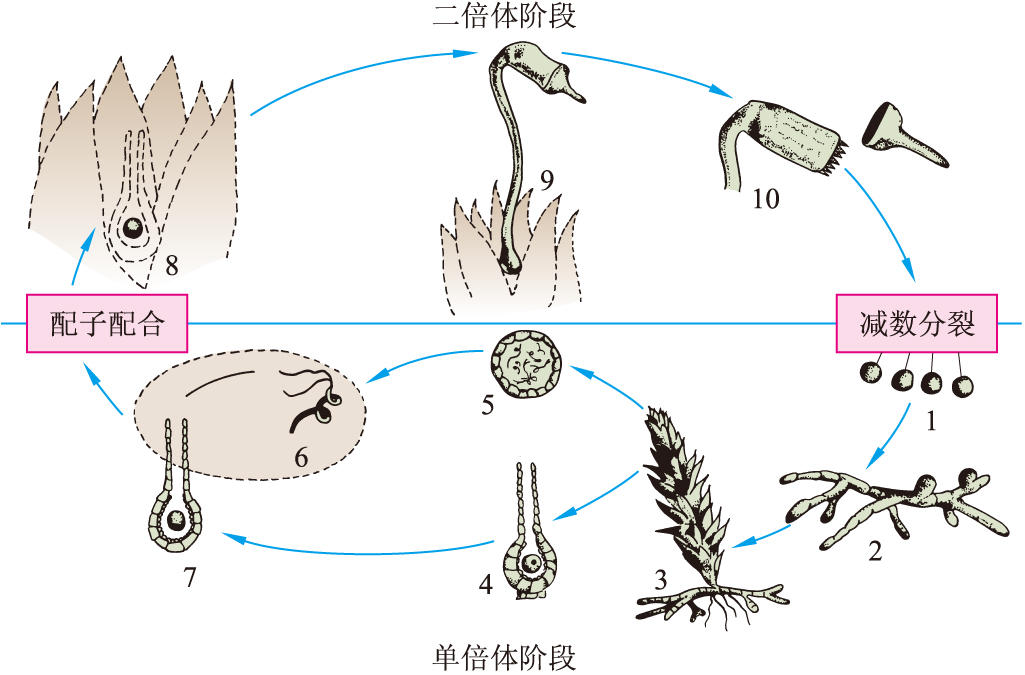 圖5 苔蘚植物不等世代型生活史(孢子體小於配子體)
圖5 苔蘚植物不等世代型生活史(孢子體小於配子體)
1 孢子微小,靠風力傳播 2 原絲體 3 配子體由原絲體上產生 4 頸卵器縱切面 5 精子器橫切面 6 雙鞭毛的精子 7 受精期,精子將從頸卵器口遊泳進入器內,與卵受精 8 合子留在頸卵器底部 9 孢子體寄生在配子體頂部 10 蘚蒴,示蘚帽已脫落,露出蘚齒,蒴內的孢子母細胞經減數分裂產生孢子
 圖6 海帶不等世代型生活史(孢子體大於配子體)
圖6 海帶不等世代型生活史(孢子體大於配子體)
1 遊泳孢子 2~5 雌配子體,隻有一卵囊和一個卵 6~8 雄配子體 9 精子 10 合子 11~15 合子萌發小孢子體 16 孢子體 17 孢子體橫切面示遊泳孢子囊
 圖7 蕨類植物不等世代型生活史
圖7 蕨類植物不等世代型生活史
1 孢子微小,靠風傳播,能漂遊空氣中 2 原葉體,示綠色組織上部為頸卵器,下部為精子器和假根 3 頸卵器縱切面 4 精子器縱切面 5 精子 6 合子留在頸卵器底部 7 小孢子體由頸卵器長出 8 已成長的孢子體,具根、莖、葉 9 葉的背面生長孢子囊群
 圖8 裸子植物(松)不等世代型生活史
圖8 裸子植物(松)不等世代型生活史
1~3 孢子體幼苗期 4 雄球果 5 雄球果直切面,示小孢子葉和小孢子囊 6 小孢子母細胞 7 四個小孢子由小孢子母細胞經過減數分裂形成 8 小孢子(花粉粒) 9~11 雄配子體的發育過程,示花粉管、精核和管核 12 雌球果幼期 13 雌球果直切面,示大孢子葉和大孢子囊在胚珠中 14 大孢子囊直切面,示大孢子母細胞 15 四個大孢子,由大孢子母細胞經過減數分裂形成 16~18 雌配子體的發育過程,示自由核、周壁和胚乳 19~21 頸卵器發育過程 22 受精期,示花粉管(雄配子體)經大孢子囊與頸卵器中卵細胞受精 23~29 精核與卵核配合成為合子,合子發育形成胚胎 30 種子直切面,示種皮、胚乳、胚
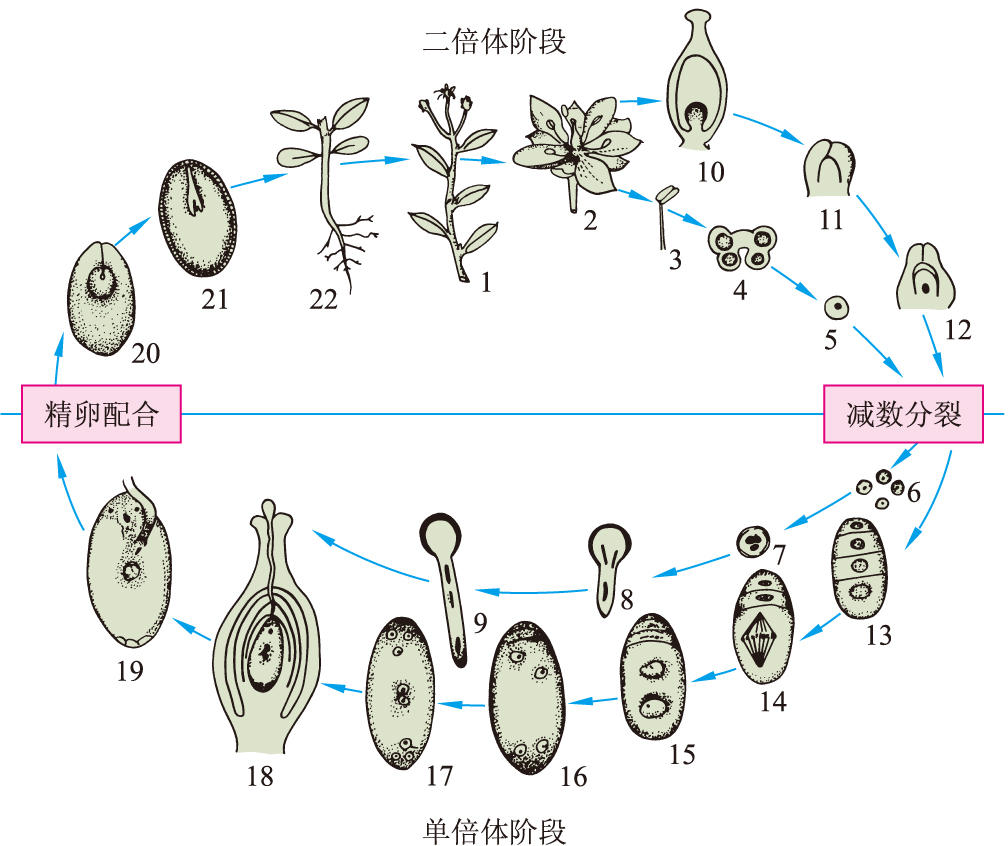 圖9 被子植物不等世代型生活史
圖9 被子植物不等世代型生活史
1 孢子體 2 花 3 雄蕊 4 花藥橫切面 5 花粉母細胞(小孢子母細胞) 6 花粉(小孢子) 7~9 花粉萌發花粉管,表示兩個精核和一個管核 10 雌蕊直切面 11 胚珠 12 胚珠產生胚囊母細胞(大孢子母細胞) 13~17 胚囊(大孢子)的發育 18 雌蕊直切面,示花粉管伸入胚囊 19 花粉管放出兩精核和一管核到胚囊 20 胚和胚乳的發育 21 成熟的種子 22 幼苗
起源與演化 藻類和菌類等低等植物的生活史類型比較多樣化,而苔蘚、蕨類以及種子植物的生活史中隻有雙單倍體類型,因而比較一致。法國人A.帕舍爾等發現幾種衣藻 (Chlamydomonas pertusa,C.Paradoxa,C.botryedes)的合子常常變成特殊的形狀,並能保持10天以上的遊動能力,形同獨立的個體。因此認為衣藻可能也有二倍體的個體。帕舍爾還發現鹽藻(Dunaliella)的合子發芽時,增加鹽分的濃度,就會長成一個至兩個具有四根鞭毛的個體。因此認為世代交替的現象,可受外界環境條件的幹預而改變,合子很有可能在某種情況下長成幾倍體的新個體。
英國學者F.E.弗裡奇根據以上研究,於1956年提出單體型、單倍體的藻類植物可能起源於能夠遊動的單細胞的綠藻,這是所有的生活史中最原始的一類。這一類藻類植物的合子,延長瞭生活期,就產生瞭單體型雙倍體的種類,若是合子經過突變,就會發展出雙單體型等世代的種類。等世代的雙單體型種類,配子體再高度簡化,就發展成雙體型不等世代的種類。
理論與實踐意義 植物生活史的研究,對於古植物學和植物分類學的發展有很大的推動作用。例如銀杏和蘇鐵的分類地位長期沒有得到肯定。直到19世紀末,日本學者平瀨作五郎和池野成一郎分別研究瞭它們的生活史的各個階段,發現它們的生殖過程還保留著具纖毛的精子,由此才確定瞭它們都是孑遺的裸子植物。
在實踐方面,如過去(1865)把小麥稈銹病菌的三種孢子,分別定為復孢銹屬、雙孢銹屬和銹孢屬三個獨立的屬;後經德國著名植物學傢H.A.德巴裡精細研究生活史才確定瞭三者是小麥稈銹病菌生活史上的三個不同階段,從而為防治這種病菌的傳播提供瞭重要的科學依據。又如紫菜屬的生活史上有一個絲狀體階段,生長在文蛤等的貝殼裡,長期被視為一種紅藻──殼斑藻。經過中國藻類學傢曾呈奎等的研究,證明它不過是紫菜生活史上的一個階段,糾正瞭過去分類上的錯誤,同時又利用這種孢子,為生產上解決瞭紫菜葉狀體人工養殖的孢子來源問題。海帶屬原是北溫帶的海藻,也是經過中國藻類學傢曾呈奎等對其生活史的研究,突破瞭孢子放散生長成小海帶的條件,使海帶的人工養殖,可從北溫帶南移到亞熱帶海岸線放養。